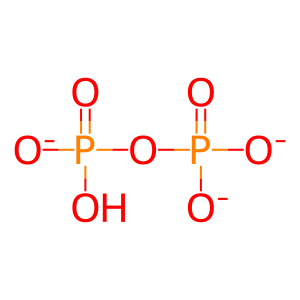
Reaction: POLI incorporates dNMP opposite to damaged DNA base
Hoogsteen base pairing and rotation of template purines from anti to syn conformation serves as a mechanism to displace adducts on template G or template A that interfere with DNA replication, as is the case with gamma-hydroxy-1,N2-propano-2'deoxyguanosine (gamma-HOPdG), or to allow base pairing of damaged purines with a disrupted Watson-Crick edge but an intact Hoogsteen edge, as is the case with 1,N6-ethenodeoxyadenosine (EtAD) (Nair et al. 2006).
Gamma-HOPdG is formed when acrolein, an alpha,beta-unsaturated aldehyde generated as an end product of lipid peroxidation or oxidation of polyamines, reacts with the N2 of guanine, leading to formation of a cyclic adduct. POLI incorporates dCMP opposite gamma-HOPdG as efficiently as opposite an undamaged G (Washington et al. 2004). EtAD is generated when DNA is exposed to chemical carcinogens, such as vinyl chloride, or epoxyaldehydes that are produced by lipid peroxidation. POLI shows preferential dTMP incorporation opposite to a template EtAD, but can also erroneously incorporate dCMP although with four times lower efficiency (Nair et al. 2006).
======