Reaction: Prolyl 4-hydroxylase converts collagen prolines to 4-hydroxyprolines
- in pathway: Collagen biosynthesis and modifying enzymes
Collagen was for many years considered the only source of 4-hydroxyproline (4-Hyp) in animals. Though it is now known that other proteins such as C1q and elastin also contain 4-Hyp, collagen is by far the major source (Adams & Frank 1980). 4-Hyp is required for collagen stability at physiological temperatures. The abundance of Hyp in animal proteins is ~4%, making it more abundant than the amino-acids Cys, Gln, His, Met, Phe, Trp and Tyr (McCaldon & Argos 1988). In collagen Hyp abundance is much higher at ~38% (Ramshaw et al. 1998). Full collagen proline hydroxylation significantly raises the melting temperature (Tm) by stabilizing the collagen triple helix (Berg & Prockop 1973a), a process that has been studied extensively using synthetic collagen peptides (Sakakibara et al. 1973, Holmgren et al. 1998) and is well understood at the structural level (Shoulders & Raines 2009). Collagen 4-Hyp content is relatively stable, with small differences between collagen types. Collagen type I has approximately 1 4-Hyp for every 10 residues, roughly 50% of available prolines (Kivirikko et al. 1992). Conversion of Pro to (2S,4R)-4-hydroxyproline (4-Hyp) is the most prevalent posttranslational modification in humans, catalyzed by prolyl 4-hydroxylase (P4H). Mammalian prolyl 4-hydroxylase is an alpha2 beta2 tetramer (Berg & Prockop 1973b). The 59-kDa alpha subunit contains the substrate-binding domain and the enzymic active site (Helaakoski et al. 1989). Humans and most other vertebrates have three isoforms of the alpha subunit, isoform alpha-1 is the most prevalent. The pair of alpha subunits can be any of the three isoforms (Gorres & Raines 2010). The 55-kDa beta subunit is protein disulphide isomerase (PDI), which has additional functions in collagen formation. As part of P4H it retains the tetramer in the ER lumen and maintains the otherwise insoluble alpha subunit in an active form (Vuori et al. 1992, Nietfeld & Kemp 1981). P4H is a member of the non-heme iron(II), alpha-ketoglutarate-dependent dioxygenase family. Molecular oxygen (O2), 2-oxoglutarate (alpha-ketoglutarate) and iron(II) are required for its activity (Hutton & Udenfriend 1966). During the reaction, alpha-ketoglutarate is oxidatively decarboxylated producing succinate and CO2 (Rhoads & Udenfriend 1968, Gorres & Raines 2010). Ascorbate is required as a cofactor but not consumed (Kivirikko et al 1989). The minimum substrate required for hydroxylation is an Xaa-Pro-Gly tripeptide, with Pro preferred in the Xaa position, though hydroxylation can occur at lower rates with a variety of residues at this position (Kivirikko et al. 1972). A number of other peptides, notably elastin, are substrates for P4H (Bhatnagar 1978).
For brevity, all forms of collagen propeptide are shown as having 3X 4-Hyp residues following the action of P4H.
For brevity, all forms of collagen propeptide are shown as having 3X 4-Hyp residues following the action of P4H.
Reaction - small molecule participants:
SUCCA [endoplasmic reticulum lumen]
CO2 [endoplasmic reticulum lumen]
O2 [endoplasmic reticulum lumen]
2OG [endoplasmic reticulum lumen]
Reactome.org reaction link: R-HSA-1650808
======
Reaction input - small molecules:

dioxygen
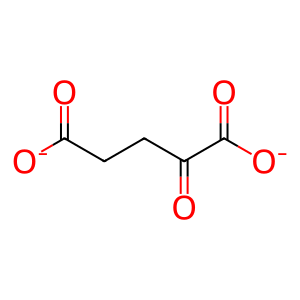
2-oxoglutarate(2-)
Reaction output - small molecules:
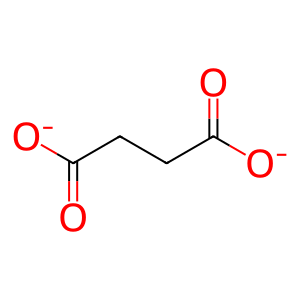
succinate(2-)
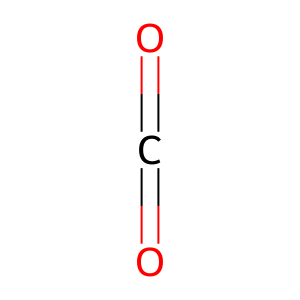
carbon dioxide
Reactome.org link: R-HSA-1650808